
托马斯·邓肯博士
语言
WEB资源
RESEARCH PROGRAMS AND AFFILIATIONS
研究兴趣
F-type ATP合酶s; bioenergetics of pathogenic bacteria; enzymology; structural biology, 膜蛋白功能.
教育利益
Bioenergetics; mitochondria; lipid and cholesterol metabolism
协会/会员
教育
研究抽象
My research focuses on the structure and function of the F-type ATP合酶, 一种能量转导酶复合物,对大多数生物的能量代谢至关重要. 我目前的重点是它在细菌代谢,毒力和发病机制中的关键作用. F-type ATP合酶 is assembled on the inner cell membrane of bacteria, 真核生物的线粒体内膜和叶绿体的类囊体膜(见“简单”细菌酶的例子), 无花果. 1). In the terminal step of oxidative- and photo-phosphorylation, it uses energy from a transmembrane, electrochemical gradient of protons (proton-motive force, or 及) to drive the synthesis of most cellular ATP. 然而,这种酶在本质上是可逆的,如果失去了呼吸的驱动力(如.g.,缺乏2)或光合作用(黑暗),酶可以转变为水解细胞ATP,驱动质子(H+) 运输 across the membrane in the opposite direction and generating 及. 该酶对各种致病菌的生存能力和/或毒力至关重要, 其中一些依靠它来合成ATP,而另一些则只能将它作为ATP驱动的H+ 泵. 尽管细菌和真核生物形式的酶在结构/功能上普遍保持一致, 贝达喹啉的开发证实了靶向这种酶攻击致病菌的潜力, which was found to target mycobacterial FOF1 and is currently approved as part of a 3-drug, front-line regimen to treat multi-drug resistant Tuberculosis. 贝达喹啉是一种新的二芳基喹啉类抗生素中的第一种,对F分枝杆菌具有高度选择性OF1.

无花果. 1. E的漫画. 杆菌FOF1 体系结构. Arrow below the c-ring shows direction the rotor complex (c10γε) turns during synthesis of ATP, as driven by H+ 运输.
BACKGROUND
ATP合酶由两个功能不同但耦合的亚复合物组成. The basic architecture of the enzyme is shown in cartoon form in 无花果. 1 for the minimal subunit composition found in most bacteria. FO 络合物催化H+ 运输 and has subunits that span the membrane bilayer. F1 是一种延伸到水相的外周络合物,包含三个催化核苷酸结合位点,用于ATP的合成和水解. FO 和F1 是通过两个茎状连接的子单元耦合的:一个中央转子轴和一个外围定子. 我和理查德·克罗斯的早期研究提供了关键的初步证据证明ATP合酶的能量耦合涉及
- Rotation of the central shaft within F1 协调三个合作的,交替的催化核苷酸位点的作用,和
- F内亚基的旋转O during energy-driven proton 运输.
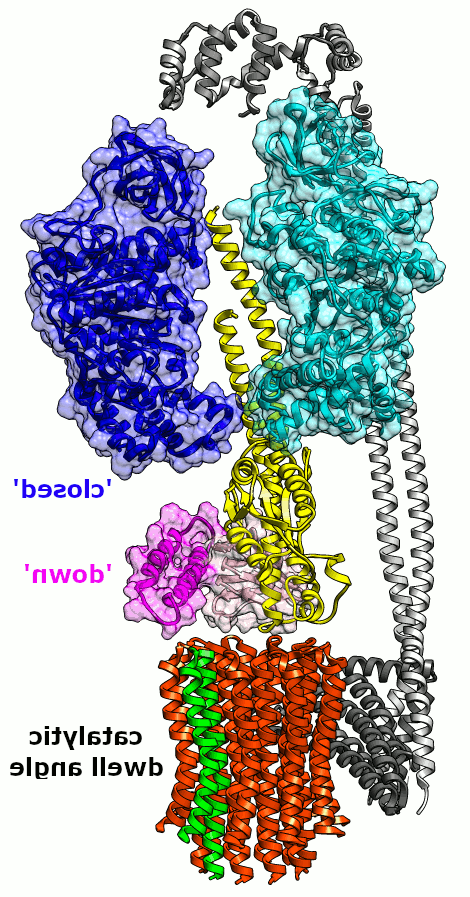
无花果. 2. 最小的 E. 杆菌 FOF1 动画. 一个β亚基的可能构象转变和相对于一个小(~36°)旋转子步的εCTD. One c-subunit is green to track rotation of the c-ring along with the rotor shaft of γ (yellow) and ε (NTD/CTD, pink/magenta). 只有1个β(青色)被显示出来,而中间的α亚基没有显示出来.
实验系统 & 当前的项目
Our research has long used the bacterial ATP合酶 from E. 杆菌, 这为这种旋转运动酶的结构/功能研究提供了一个简单而有力的模型. E. 大肠杆菌系统允许直接的遗传筛选和ATP合成酶的工程, and also large-scale purification of FOF1 分离的可溶性F1 for biochemical and structural studies. We continue to develop more detailed knowledge of E. 杆菌 FOF1 了解εCTD的抑制机制,以及如何更好地靶向它来破坏细菌的生物能量学. For example, an ongoing collaboration with a single-molecule microscopist, Dr. 迈克尔·罗宋汤 (耶拿大学, 德国)利用荧光共振能量转移(FRET)研究了ε在单个EcF中的构象变化动力学OF1脂质体.
我们对F的了解要少得多OF1 我们已经开始研究FOF1 在两种不同的病原体中:
- 铜绿假单胞菌. 这革兰氏阴性, 机会致病菌是囊性纤维化患者持续肺部感染和死亡的主要原因. 它对免疫功能低下的人是一种重大危害,在医院获得性感染中很普遍. Multi-drug-resistant strains continue to emerge and, in 2017, the World Health Organization listed carbapenem-resistant P. 绿脓杆菌 as one of three pathogens in critical need of new antibiotic development. P. 绿脓杆菌 新陈代谢强烈依赖于呼吸,ATP合酶对其生存能力至关重要. We are currently collaborating with the labs of 克里斯托弗。野村 在邻近的SUNY-ESF和 Guirong王 (推荐最近最火的赌博软件 Dept of Surgery) to study (i) how the εCTD of FOF1 impacts the enzyme’s function in P. 铜绿菌体外培养 (ii) εCTD的破坏是否影响病毒的毒力或发病机制 P. 绿脓杆菌 in cultured macrophages and in a mouse model of lung infection.
- 链球菌引起的肺炎. 这种革兰氏阳性病原体是细菌性肺炎的主要原因,但也可导致严重的侵袭性肺炎球菌疾病,如败血症和脑膜炎. 这种耐氧厌氧菌缺乏完整的呼吸链,因此不能利用FOF1 重要的ATP合成. 然而,FOF1 是必不可少的 S. 肺炎的 viability and acid tolerance, acting as an ATPase-driven H+ 泵通过细胞膜产生及并维持细胞pH稳态. A recent study of septicemia in mice, by the research group of 马可Oggioni (英国莱斯特大学)的研究表明,肺炎球菌F具有重要的毒力作用OF1: S的亚克隆. 肺炎,出现建立败血症最常包含一个新生的错义突变基因的一个亚单位的FOF1 (2014, Gerlini et al ..中国生物医学工程学报(英文版). One such virulent clone instead had a frame-shift mutation in the atpC gene for ε that ‘scrambled’ the sequence of the εCTD, and that pneumococcal clone showed increased survival in the spleen. 他们转移了 atpC 突变成naïve, nonencapsulated pneumococcal strain, 我们正在研究εCTD的破坏是如何影响F的OF1 function in pneumococcal membranes.
选择引用
T.M. 邓肯 (2019). Turbine enzyme’s structure in the crosshairs to target tuberculosis. Proc. 国家的. 阿德莱德大学. Sci. 美国 116, 3956-58. http://doi.org/10.1073/pnas.1900798116
M. 索R. Ishmukhametov J.C. bouw,. 昨天,C. Suarna N.J. 史密斯,米. 克里斯蒂,R. 装料工T.M. 邓肯, A.G. 斯图尔特(2019). Cryo-EM reveals distinct conformations of E. 杆菌 ATP合酶对ATP的影响. eLife 8, e43864. http://doi.org/10.7554/eLife.43864
H. Sielaff T.M. 邓肯, M. 罗宋汤(2018). 调节亚基ε in 大肠杆菌 FOF1atp合酶. Biochim. Biophys. Acta Bioenerg. 1859, 775-788. http://doi.org/10.1016/j.bbabio.2018.06.013
N.B. 沙,T.M. 邓肯 (2015). 好氧生长 大肠杆菌 当从ATP合酶的ε亚基上删除五个c端残基时,ATP合成被选择性地抑制, J. 医学杂志. 化学. 290, 21032-41. http://doi.org/10.1074/jbc.M115.665059
M. 罗宋汤,T.M. 邓肯 (2013). Spotlighting motors and controls of single FOF1atp合酶. 物化学. Soc. 反式. 41, 1219-26. http://doi.org/10.1042/BST20130101
N.B. 沙,M.L. 哈钦,B.K. 哈雾,T.M. 邓肯 (2013). F1腺苷三磷酸酶的 大肠杆菌: The ε-Inhibited State Forms After ATP Hydrolysis, is Distinct from the ADP-Inhibited State, and Responds Dynamically to Catalytic-Site Ligands. J. 医学杂志. 化学. 288, 9383-95. http://doi.org/10.1074/jbc.M113.451583
G. Cingolani T.M. 邓肯 (2011). Structure of the ATP合酶 catalytic complex (F1) 大肠杆菌 in an autoinhibited conformation. Nat. Struc. 摩尔. 医学杂志. 18, 701-7. http://doi.org/10.1038/nsmb.2058
T.M. 邓肯 (2004). The ATP Synthase: Parts and Properties of a Rotary Motor, in: 酶,卷. XXIII: Energy Coupling and 摩尔ecular Motors,卷. 23, (D.D. 哈克尼F. Tamanoi, Eds.) Elsevier 阿德莱德大学emic Press, New York, pp. 203-275.
M.L. 哈钦T.M. 邓肯, H. Ngai, R.L. 十字架(2001). Energy-driven subunit rotation at the interface between subunit a 和 c F的低聚物O 部门的 大肠杆菌 ATP合酶. Proc. 国家的. 阿德莱德大学. Sci.. 美国 98, 8519-24. http://doi.org/10.1073/pnas.151236798
T.M. 邓肯, V.V. Bulygin Y. 周,M.L. 哈钦,R.L. 十字架(1995). Rotation of subunits during catalysis by 大肠杆菌 F1腺苷三磷酸酶. Proc. 国家的. 阿德莱德大学. Sci. 美国 92, 10964-68. http://doi.org/10.1073/pnas.92.24.10964